
The SIR model was a demonstration designed to make a point
It was not meant for general disease modeling

I work in infectious disease modeling, and I am often asked if I use “SIR” models, that is, systems of ordinary differential equations that model infection as three states: susceptible, infectious, and recovered.
The short answer is no, I don’t. Rather than explain why not, I’d rather explain why the SIR model came to be in the first place.
The SIR model was not intended, as most people expect, to simulate any particular outbreak of any particular disease. Instead, the SIR model was a mathematical construct designed to prove an important but very general point about the dynamics of infectious disease outbreaks.
The SIR model comes from a famous 1927 paper by the mathematical epidemiologists Kermack and McKendrick. They make clear they are trying to solve a fundamental problem in epidemiology. Why doesn’t every disease outbreak, or “epidemic,” infect everyone on the planet? In an outbreak, the number of newly infected people starts small, goes up to a peak, and then comes back down. Why?
One of the most striking features in the study of epidemics is the difficulty of finding a causal factor which appears to be adequate to account for the magnitude of the frequent epidemics of disease which visit almost every population. It was with a view to obtaining more insight regarding the effects of the various factors which govern the spread of contagious epidemics that the present investigation was undertaken. [...] The problem may be summarised as follows: One (or more) infected person is introduced into a community of individuals [...] The disease spreads from the affected to the unaffected by contact infection [i.e., directly from human to human, not via a polluted water source, mosquito, etc.]. Each infected person runs through the course of his sickness, and finally is removed from the number of those who are sick, by recovery or by death. [...] As the epidemic spreads, the number of unaffected members of the community becomes reduced. [...] In the course of time the epidemic may come to an end. One of the most important problems in epidemiology is to ascertain whether this termination occurs only when no susceptible individuals are left, or whether the interplay of the various factors of infectivity, recovery and mortality, may result in termination, whilst many susceptible individuals are still present in the unaffected population.
They set out to show that the two leading explanations of the day were either unnecessary or incomplete:
Two of the reasons commonly put forward as accounting for the termination of an epidemic, are (1) that the susceptible individuals have all been removed, and (2) that during the course of the epidemic the virulence of the causative organism has gradually decreased.
In other words, there were two leading hypotheses. First, the outbreak stops because everyone who could be infected was infected. This theory would have presented difficulty, even with the data available at the time. For example, it was well-known that measles, before the introduction of the measles vaccine, followed a two-year cycle: a year in which many children were infected was followed by a year with fewer infections. From this it would be clear that there were some children who didn’t get measles one year but who got it the next, which doesn’t fit with the hypothesis.
The second leading hypothesis was that, in every outbreak, the pathogen causing the outbreak begins as very capable of spreading between people and causing infected people to become sick, and then slowly loses that power. This idea was about as old as the germ theory of disease. Jacob Henle, in his 1840 On Miasmata and Contagia, tried to reason about this idea, based on the muddled and contradictory data available to him:
Pessina suggested a means of attenuating the intensity of the contagion, as follows: that ten sheep be inoculated from a sheep with natural pocks, that then others again be inoculated from that one of the first group which has the best pocks, etc. By this continued involution, as a homeopath would call it, the pox should become continually milder and produce fewer vesicles, until the eruption finally confines itself to the single point of inoculation.
This claim is opposed by more recent practitioners. The protective power and the milder course of the vaccinated sheep-pox has been put beyond a doubt, namely by Kaush and Müller, and Krüger, who at the same time also showed that the cultivated pox doesn’t at all lose its intensity as continually, as the Austrian authors claim, and that general eruptions are rather frequent, irregularly intermittent, and appear again in later generations.
The idea that outbreaks must come down because pathogens must become milder hasn’t died out; there was a common but misguided belief that every new COVID-19 variant would be less dangerous than the last.
Kermack and McKendrick proposed a different hypothesis to explain the finite size of outbreaks:
It will be shown [...] that [...] the course of an epidemic is not necessarily terminated by the exhaustion of the susceptible members of the community. It will appear that for each particular set of infectivity, recovery and death rates, there exists a critical or threshold density of population. If the actual population density be equal to (or below) this threshold value, the introduction of one (or more) infected person does not give rise to an epidemic, whereas if the population be only slightly more dense, a small epidemic occurs. [... T]he epidemic continues to increase so long as the density of the unaffected [i.e., susceptible] population is greater than the threshold density, but when this critical point is approximately reached, the epidemic begins to wane, and ultimately to die out. This point may be reached when only a small proportion of the susceptible members of the community have been affected.
Kermack and McKendrick pose their theory in terms of “density,” but I find it easier to think about rates in time: above the epidemic threshold, the rate of new infections is greater than the rate at which infectious people cease being infectious, and the outbreak grows. Below this threshold, infectious people recover faster than they produce more infections, and the outbreak shrinks. (In epidemiology jargon, the “effective reproduction number” falls below one.)
In short, they hypothesize that outbreaks (of this type, at least) do not cease because there are no more susceptible people, but because each infectious person cannot infect enough new people to keep the outbreak growing.
Rather than follow Kermack’s and McKendrick’s original derivation, which can be difficult for a modern reader to follow, I’ll lay out my own argument, arriving at the same conclusion. Our goal is to build a model that satisfies these requirements:
The number of susceptibles does not go to zero at the end of the outbreak.
The properties of the pathogen do not change during the outbreak.
Some interplay between the number of susceptibles, infectivity, and recovery leads to the end of the outbreak.
The model can be built with the knowledge and tools available in 1927.
First, to avoid depleting the number of susceptibles, let us consider an infinite population. This way, not only will the number of susceptibles not go to zero at the end of the outbreak, it will in fact be infinite.
Second, assume that the population is perfectly connected: every person in this infinite soup of people has an equal opportunity to interact with every single other person. Clearly this is impossible, and not a good modeling assumption for any actual outbreak, but we are here to prove a point, and I want to avoid any claim that somehow our model is causing a “local” depletion of susceptibles. Let’s make this impossible by assuming that any single infectious person is legion, that they could, in theory, infect everyone in this infinite universe of people.
Third, we say that each infectious person’s ability to infect more people is drawn from the same distribution, which remains constant over the outbreak. Each infectious person need not produce exactly the same number of onward infections, but they must have the same a priori capability to do so.
Fourth, in order to make the model tractable with 1927 technology, we assume all susceptible people identical. In reality, different people would be differently susceptible to the disease in question. For the purposes of this toy model, we make the simplifying assumption that all susceptible people are equally susceptible. This means we don’t need to keep track of each susceptible person, but instead can summarize the susceptibles by a single number, the proportion of all people in the infinite population who remain susceptible.
Fifth, we must make an analogous assumption for infectious people, so that we can track this infinite population of infectious people with a finite set of numbers, ideally just one. We can easily assume that all infectious people are equally “infectious,” that is, capable of infecting the same number of other people per unit time.
Naively, one might also expect we can assume that every infected person is infectious for the same, fixed amount of time. In fact, this requires keeping track of the time each infected person became infected, which was impossible for 1927 technology. Today I can easily run a computer simulation like this with one million people, but Kermack and McKendrick could only have done this for a very small number of people, which would not have produced a model that would prove their point.
In order to make all infectious people “the same,” each infectious person must have the same chance to stop being infectious, at any given moment, regardless of when they were infected. Whether you have been infected for one second, one day, or one month, your chance of recovering from infection in the next second is the same. (In statistics jargon, this is called “memorylessness.”) Mathematically, this assumption requires that the distribution of infectious periods be exponential, meaning that most people have very short infectious periods, a few have medium periods, and a vanishingly small number have very long infectious periods.
This would clearly be a bad assumption for any real disease outbreak. For most diseases, most people have “medium” infectious periods. I can’t think of any disease that has anything like exponentially distributed infectious periods. At this point, one can begin apologizing, creating intermediate disease states so that the distribution of infectious periods is not exponential but something more biologically plausible. Instead, let us remember that this is a toy model, designed to prove a particular point. (We could use those intermediate disease states to relax our core assumptions, and show that the key result about the existence of an epidemic threshold does not depend on the precise exponential distribution assumption, but that is a tactic to improve a demonstration, not to create a verisimilitudinous model.)
These assumptions —infinite population, well-connected population, identical susceptibility, identical infectiousness per unit time, exponentially-distributed infectious periods— together produce the familiar SIR differential equations, which in turn display Kermack’s and McKendrick’s desired behavior: there is a threshold combination of proportion susceptible, and infection rate. Above this threshold, an epidemic grows; below, it shrinks. In every parameter combination, at the end of the outbreak, there will always be some nonzero proportion of people who remain susceptible. That fraction might be very small, but it is never zero.1
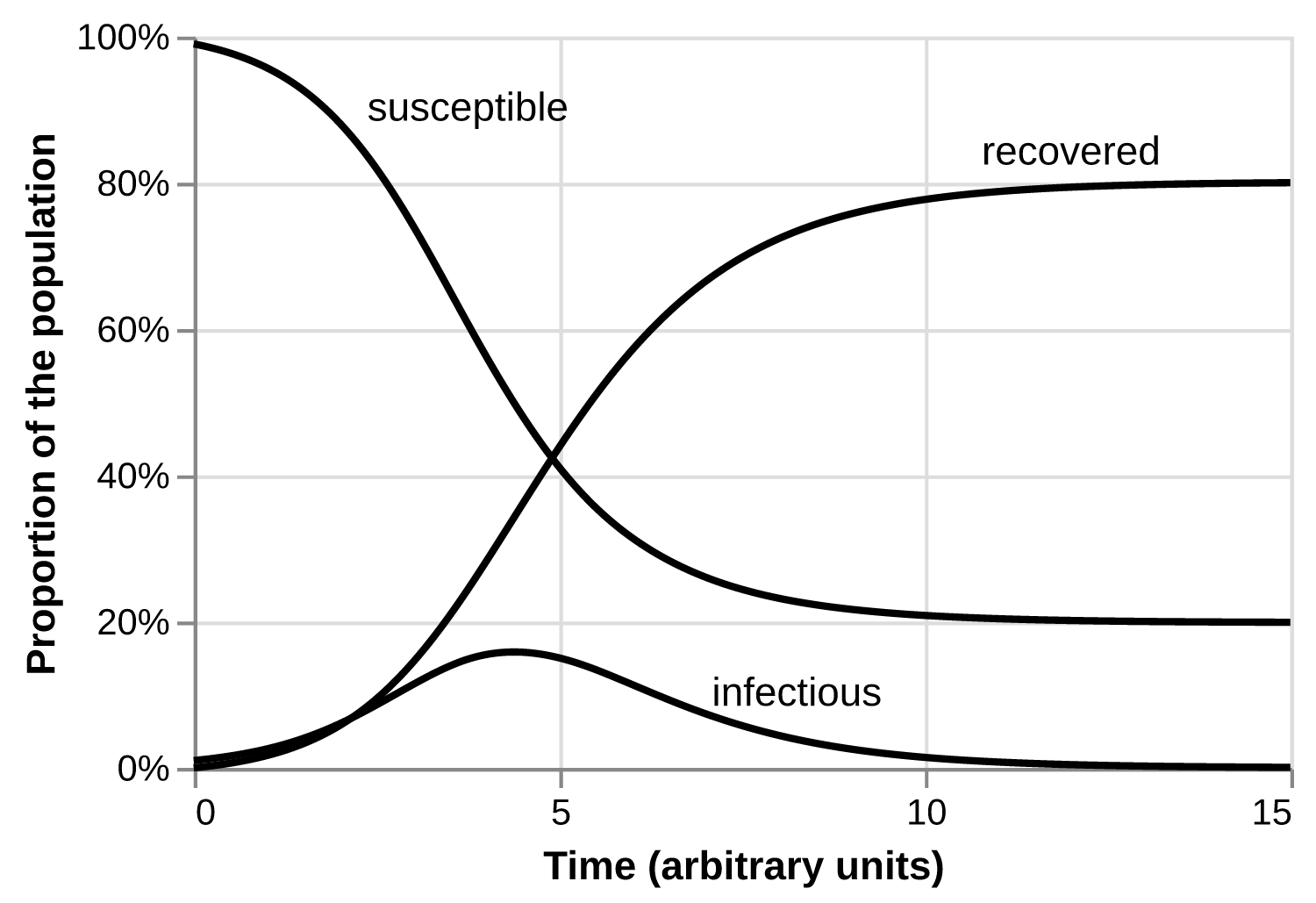
The SIR model unraveled a long-running mystery in epidemiology: an outbreak of a disease, in a perfectly connected population, transmitted from person-to-person, whose infection bestows lifelong immunity, does not need to infect everyone before the outbreak stops. Extensions of the model show what happens when the duration of infectiousness is not exponentially distributed, or if you allow births, or if you have different classes of susceptibles, or if you have two competing diseases, or if immunity is not permanent, or any number of other questions.
Mathematically, the SIR model is really wonderful: its simplest form is amenable to undergraduate mathematical analysis, but it has just enough complexity to address some interesting, fundamental questions in epidemiology.
The model’s success in its original purpose, and its mathematical tractability, have also led to its abuse. It is a theoretical proof of a general pattern, not a law of nature. The assumption of an infinite population leads to weird effects. For example, outbreaks reach a finite size, but the number of infectious people never reaches zero. And while SIR dynamics lead to outbreaks of finite size, the fact that a particular outbreak goes up and comes down does not mean that reality is conforming to SIR dynamics. Foodborne outbreaks go up and come down for reasons unrelated to the depletion of susceptibles, and the waves of COVID-19 were driven by a combination of changing human behavior and an evolving pathogen.
Like all tools, the SIR model was designed for a purpose. It’s just not the purpose most people know.
Unless you start with zero susceptibles or zero infectivity or something else smart-alecky.